- Новости науки и техники
- Биология и медицина


31 октября 2025, 07:27
Аппетит как мишень: почему все попытки обуздать ожирение и диабет проваливались, пока не появились агонисты рецепторов инкретинов
Ожирение и сахарный диабет 2 типа приобрели масштабы глобальной пандемии и уже успели создать беспрецедентную нагрузку на системы здравоохранения.
Читать ещё
31 октября 2025, 02:19
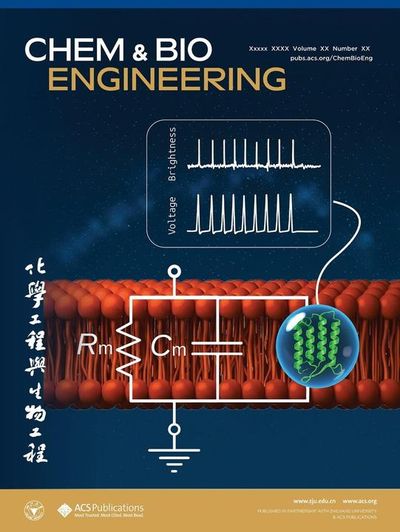
Созданы более яркие белки–сенсоры для исследования активности головного мозга и сердца
Ученые разработали светящиеся молекулярные сенсоры для наблюдения за активностью живых клеток, которые значительно ярче и чувствительнее аналогов. Такие молекулярные датчики позволяют отслеживать электрические сигналы в клетках, благодаря чему могут использоваться при изучении работы мозга и сердца с помощью флуоресцентного микроскопа
Читать ещё
31 октября 2025, 02:19
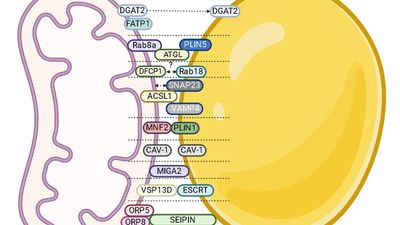
Ученые нашли потенциальные мишени для борьбы с метаболическим синдромом
Ученые БФУ имени Иммануила Канта проанализировали механизмы развития метаболического синдрома — нарушения обмена веществ, которое приводит к ожирению, повышению артериального давления, высокому уровню сахара и холестерина в крови. Оказалось, что потенциальными мишенями для лечения этого состояния могут стать белки, отвечающие за взаимодействие митохондрий и липидных капель в клетках, поскольку нарушение их работы приводит к инсулинорезистентности и избыточному накоплению жиров в организме
Читать ещё

Ученые обнаружили переключатель, который может ослаблять и усиливать воспоминания. Революционное открытие предлагает новые возможности для лечения ПСТР, зависимостей и деменции.
Читать ещё

В Калифорнийском университете предлагают лечить генерализованное тревожное расстройство с помощью фармацевтической формы ЛСД — препарата MM120. Клинические исследования показали, что однократная доза препарата куда лучше стандартных антидепрессантов облегчает симптомы у пациентов с хронической тревогой. Это достигается за счет стимуляции нейропластичности и преодоления устоявшихся моделей мышления.
Читать ещё

N + 1 — главное издание о науке, технике и технологиях
Читать ещё
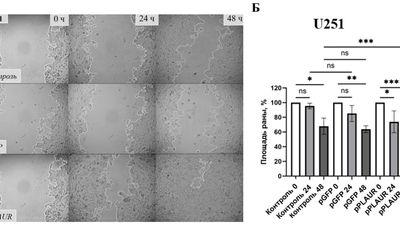
Ученые БФУ имени Иммануила Канта определили молекулярный механизм, за счет которого такие опухоли как глиомы и нейробластома становятся крайне агрессивными и устойчивыми к лечению. Оказалось, что появлению метастазов и миграции опухолевых клеток по организму способствует повышенная активность гена рецептора урокиназы PLAUR, регулирующего «перестройку» внеклеточного вещества в тканях
Читать ещё

28 октября 2025, 18:21
Новая модель белкового языка превосходит AlphaFold3 от DeepMind
Шотландские ученые использовали в необычных целях суперкомпьютер Tursa из вычислительного центра DiRAC, который обычно применяют для космических исследований. Они разработали модель языка белков, получившую название PLM-Interact. Она предсказывает взаимодействия белков и прогнозирует, какие мутации нарушат взаимодействие между ними. Предварительные тесты показывают, что PLM-Interact превосходит существующие аналоги, включая модель AlphaFold3 компании Google DeepMind. В области медицины PLM-Inter
Читать ещё
28 октября 2025, 15:22
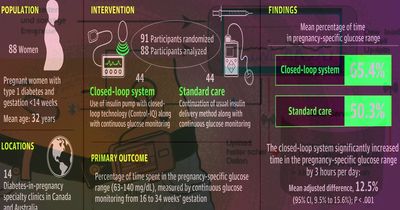
Помпы с обратной связью помогли беременным с диабетом лучше инъекций инсулина
N + 1 — главное издание о науке, технике и технологиях
Читать ещё
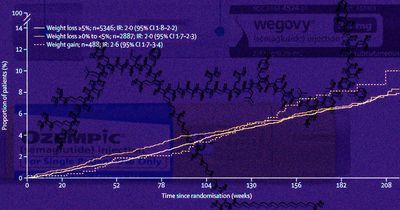
28 октября 2025, 13:26
Семаглутид снизил риск инфарктов и инсультов независимо от похудения
N + 1 — главное издание о науке, технике и технологиях
Читать ещё